 |
 |
|||||||||||||||||||||
|
||||||||||||||||||||||
JANUARY 2014 BOTTERI’S SPARROW (Peucaea botterii arizonae) SURVEYS, WITH WINTER HABITAT CONSIDERATIONS
Alan Schmierer, PO Box 626, Patagonia, AZ 85624
(All photos taken by the author)
 |
| Figure 1: Botteri’s Sparrow at location “L” from 22 January 2014 survey. Note rather fresh molt in this and subsequent photos. |
In September 2013 the author presented information reviewing the recent winter records of Botteri’s Sparrow (Peucaea botteri arizonae) in Arizona (Fig. 1; Schmierer 2013). Included in that article were the results of the author’s 6, 7 and 8 January 2013 survey for Botteri’s Sparrows along the roads of Patagonia Lake Ranch Estates, just south of Patagonia Lake, Santa Cruz Co, Arizona. Also included were the results of a February 2013 follow-up survey of a more limited area. The protocol and results of those surveys are detailed on page 4 of that article. This current publication reports the results of the author’s follow-up surveys of January 2014.
In addition to the bird survey, local botanist Sue Carnahan, assisted by the author, revisited the sites where the 2014 winter Botteri’s Sparrows had been detected to assess the vegetation species, density, height, degree of grazing, and various physical land features. A similar analysis was done at selected local sites where Botteri’s Sparrows had not been seen in either winter, comparing these to the Botteri’s detection sites.
The following information is generally presented as Part 1, describing the methodology, results and conclusions of the 2014 Botteri’s Sparrows surveys, and Part 2, discussing the methodology and results of the related habitat survey of the same area. The two considerations are then discussed as they may relate to each other.
Part 1: BIRD SURVEY
METHODS
On 8, 9, and 10 January 2014 the author did a modified, informal point-count survey that was designed and conducted to replicate the 2013 survey as close as possible. It covered the same 19.6 kilometers of roads in the development, with kilometers covered, hours, weather, and protocol being essentially the same as in 2013 (Schmierer 2013). In addition, several areas of the development where sparrows were not detected on that initial survey were re-surveyed on 22, 23, and 25 January. These additional surveys totaled 2.5 hours by car and 3.5 hours on foot, and covered 10.0 km of roads or sections of roads that had produced no detections in the winter of 2013 or 2014. The intent of the latter was to identify what areas did not appear to attract wintering Botteri’s Sparrows, for the purpose of later studying the habitat characteristics of these areas.
RESULTS
The Botteri’s Sparrow combined survey detections for 2013 totaled 25 birds at 20 locations, with a total effort of 11.7 hours; for 2014 totaled 15 birds at 12 locations, with a total effort of 12.8 hours. The total kilometers of road covered in 2014 were the same as in 2013, but in 2013 only 1.9 km were re-surveyed, compared to 10 km re-surveyed in 2014. In 2013, two incidentally found birds seen during the week were added to the totals; in 2014 one was added. This was done in order to provide the most complete picture possible. Roads surveyed and mapped locations of the 2013-2014 birds are found here.
Birds found less than 90 m from each other were considered as the “same location” and were given the same letter designation on the map. The author estimates that all marked locations are accurate to within 5 m of actual locations where each bird was first detected. The map also includes locations of several incidentally found birds. The sections of the development occupied by the 2014 birds was similar to that of the winter of 2013 and generally concentrated in the north-eastern half of the road system. Notable differences were the addition of one very southwestern area (on Lado de Loma Road) and the lack of detections on Circulo Montana. The latter includes the author’s yard, where the sparrows utilized the feeder last year but were un-detected in the winter of 2014.
Of the birds detected, only one was silent and was not detected until it was about 1.5 m from the car. All others responded to playback with high-pitched chipped call notes similar to those heard during the breeding season. The responses ranged from rather quiet chipping, but with an insistent pace, to chipping that was very strident. Distance from the playback did not seem to correlate with the intensity of the reaction. Five individuals responded with a short, emphatic trill call mixed in with the high-pitched chipping notes. Most birds, especially those with the most emphatic chipping, showed very agitated behavior, with hyperactivity, cocked tail and raised crown and nape feathers (Fig. 2 and 3).
 |
 |
| Figure 2: Botteri’s Sparrow at location “L” from 22 January 2014 survey. Note raised crown and nape feathers, in agitated reaction to playback | Figure 3: Botteri’s Sparrow at location “B” from 8 January 2014 survey. Note again the raised crown and nape feathers, in agitated reaction to playback. |
Typically these Botteri’s Sparrows responded to playback by flying from the grass up into the back side of a nearby, if not the nearest, tree or shrub, then working their way to the playback side of that tree, generally 1-2.7 m from the ground. The farthest responses to playback came from 46 m: two birds that flew to a bush 27 m away, agitated and chipping, then flew tree to tree, but never approaching closer than that 27 m. Of four other birds that responded from about 30 m, two approached and two stayed at that distance. Seven of the fifteen 2014 detections were within 90 m of a 2013 location. The longest linear distance that any bird moved while being observed was about 70 m.
No Cassin’s Sparrows (Peucaea cassinii) and only one Grasshopper Sparrow (Ammodramus savannarum) responded to the Botteri’s Sparrow playbacks.
DISCUSSION
A review of eBird, Explore Data section, revealed only two reports of Botteri’s Sparrow this winter other than those reported here: one by the author on 14 December 2013 here on Lado de Loma Road (playback used to find this species for the CBC) and one on 16 December 2013 reported by Robert Behrstock south of Sierra Vista, Cochise County (eBird 2014). This low number of sightings is not unexpected, since winter detection is generally very local and is primarily only possible using playback, a technique often discouraged by the birding community. The results of the surveys demonstrated moderate numbers present locally, but fewer than in 2013, the only reference year.
Most prior literature implied that wintering birds were silent, even in response to being flushed or to playback (Webb and Bock 2012). The most common winter response of the closely related Cassin’s Sparrow to playback was described as: “...all birds gave Chip, some flew to perch near observer, and most responded by the time 4–6 songs had been played. In these same patches, only one individual was flushed without use of the tape.” (S. M. Russell pers. comm., in Dunning et al. 1999).
In the current surveys all but one bird were vocal. The following linked audio-video, although presumably by the date and location representing a breeding bird, realistically captures the sound and action of agitated winter Botteri’s Sparrows when responding to the author’s playback: file here (recorded on 2 May 2010 by Richard Garrigues in Costa Rica).
In addition to chip notes, five birds issued an agitated trilled call, not heard by the author on any prior survey. Although all birds appeared moderately to extremely agitated, showing hyperactivity, cocking of the tail, and raised crown and nape feathers, these five birds appeared to be among the most defensive. This call sounded like one of the longer and lower pitch trill notes of the “chatter song”, the prelude to the “bouncing-ball” part of the full song.
 |
| Figure 4: Botteri’s Sparrow at location “L” from 22 January 2014 survey. This bird responded to playback with only chipping notes. |
Although both sexes issue the short chip notes as communication and alarm (Fig. 4; Webb and Bock 2012), little is known about Botteri’s Sparrow winter foraging territories or the associated vocalizations. It is known in some other passerines that non-breeding winter territories/ranges are established that may not correspond closely to breeding territories (Leahy 1982). It is further known in some species that winter in the same area where they nest that birds defend their territory throughout the year (Leahy 1982; Arcese and Sogge 2002), but that this defense may be ...”less by song and more by eviction of trespassers at close range” (Arcese and Sogge 2002). There is evidence from within-season recapture rates of wintering Cassin’s Sparrows that this closely related species is sedentary (Ruth 2000). Male winter territories of some sparrow species (e.g., Song Sparrow, Melospiza melodia) may expand 6-10 time that of breeding birds (Arcese and Sogge 2002). Territorial enforcement may be much less vigorous where migratory, stronger where resident or when in past or future breeding sites, perhaps essential toward ownership of that territory in the spring and thus assuring greater breeding success (Arcese and Sogge 2002).
The male and female of some species each have their own territory (Leahy 1982). Breeding females of some species may show aggression toward other females and that aggression may be accompanied by aggressive posture (Arcese and Sogge 2002) similar, perhaps, to that seen in some of the surveyed Botteri’s Sparows. It is clear that the Botteri’s Sparrows detected in the author’s survey show strong evidence of territorial behavior as demonstrated by strident chipping, raised feathers, and close and apparent fearless approach to the playback. This does, however, not indicate whether responding birds were male or female, as the species is not dimorphic and the vocalizations of females in winter are not well understood.
Measurement of the size of these potential winter territories was not attempted in the current surveys, nor could site fidelity (repeated winters or past/future breeding) be assessed since these were not banded birds. Botteri’s Sparrow breeding site fidelity has been found to be only moderate (25% to 43%) in this habitat (Jones and Bock 2005). That half of the 2014 birds were found near where birds were found in the 2013 winter season offers no meaningful evidence that there is any winter site fidelity because any wintering birds here would likely occupy the most favorable available habitats.
Some birds were found in close proximity (three birds at 2014 site “B” and two birds at site “A” on the map). These birds were close enough to each other that one could reasonably presume that they were all in the same territory or territory size area, yet they were silent until they responded to the playback. Several breeding males were singing close to these sites in summer 2013. Were these winter birds a family group, females or just non-territorial for other reasons? In the winter of 2013 the author found 4 similar locations where there were two birds in close proximity that showed no sign of aggression toward each other. (After the study period, on 18 March 2014, four birds were found together at site “G”. Two of these birds perched on a fence within 15 cm of each other and interacted with each other without any apparent defensive posturing. Only two of the four birds vocalized.)
PART 2: HABITAT SURVEY
METHODS
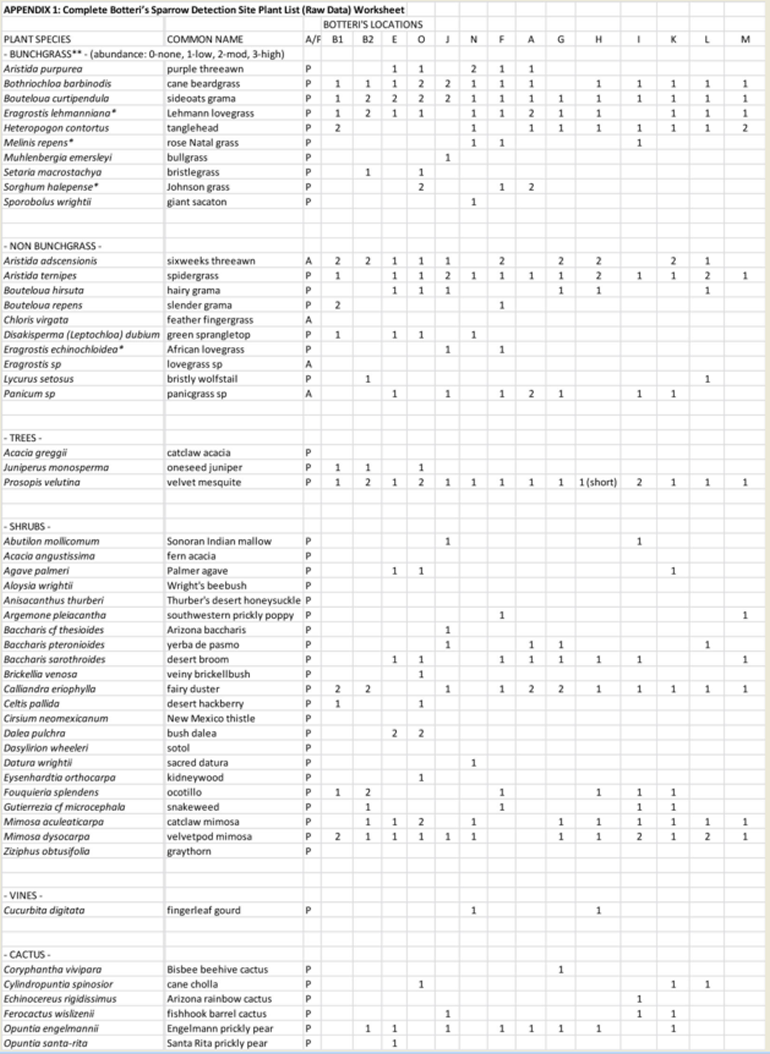
Sue Carnahan, a self-trained botanist familiar with the local flora, volunteered to assist the author by assessing the habitats of the January 2014 detected Botteri’s Sparrows. A vegetation checklist was developed by Carnahan, who then applied it on 29 January in a survey of 14 of the author’s 8-25 January 2014 surveyed Botteri’s Sparrow detection locations. On that survey Carnahan and Schmierer also noted the tree and shrub canopy, the altitude, slope angle and direction, ground vegetative cover, tallest grass, density of ground cover, and signs of recent grazing activity.
The size and nature of winter territories/winter home ranges of Botteri’s Sparrows in Arizona are unknown (Webb and Bock 2012), making any projections of knowledge of breeding territories into wintering data uncertain, at best. Furthermore, lacking a nest or singing male, as would be found in summer, establishing the center of the survey circle was equally uncertain. The 20 m radius of the survey circle used was designed to be conservative in size; small enough to be practical to survey, yet not to exceed any potential territory boundaries. The center of the circle was the location of the bird detection. If the bird moved between locations or perches, the estimated center of these locations was used. Where roads, houses or mowed yards encroached on the circle, these parts of the circle were excluded from the data. Where birds crossed the road, the study plot was split between the sides of the road.
Carnahan observed each site with the primary goal of identifying all the plant species within that 20 m circle. Toward this end, Carnahan slowly walked through each study area in approximately 3.5 m separated transects. The separation of these informal transects varied with the terrain and the ability to see each plant well enough to assure identity. The percent coverage by tree and by shrub canopies were subjectively estimated. The overall surface cover of each site was then subjectively estimated as to: percent open ground (roads, washes, ant circles, etc.), percent “bunchgrass” cover, and percent short grasses and forbs cover, where the total of these three factors equaled 100%. The tallest grass in the site was measured and recorded. The degree of grazing was estimated subjectively by Carnahan and the author and recorded as: “none”, generally fenced exclosure areas; “light” where only minor traces and/or sometimes older evidence of livestock grazing were seen; “moderate”; and “heavy”, where most of the grasses and forbs were grazed to stubble, leaving little or no cover. Finally, a subjective estimate of the overall density of cover was made. This estimate was basically a subjective assessment of the overall cover in which a sparrow could find shelter; a combination of the number of plants, percent of “bunchgrasses”, percent dense shrubs and the degree of grazing. An estimate of “0” was used to indicate bare ground and vegetation too thin and/or short to conceal a sparrow, up to a “3”, where cover was particularly thick and tall and often consisted of catclaw-type shrubs and especially thick and tall “bunchgrasses”, thus providing almost unlimited cover. At two sites where access was limited by fences or the terrain was very steep, binoculars were used to estimate the survey parameters.
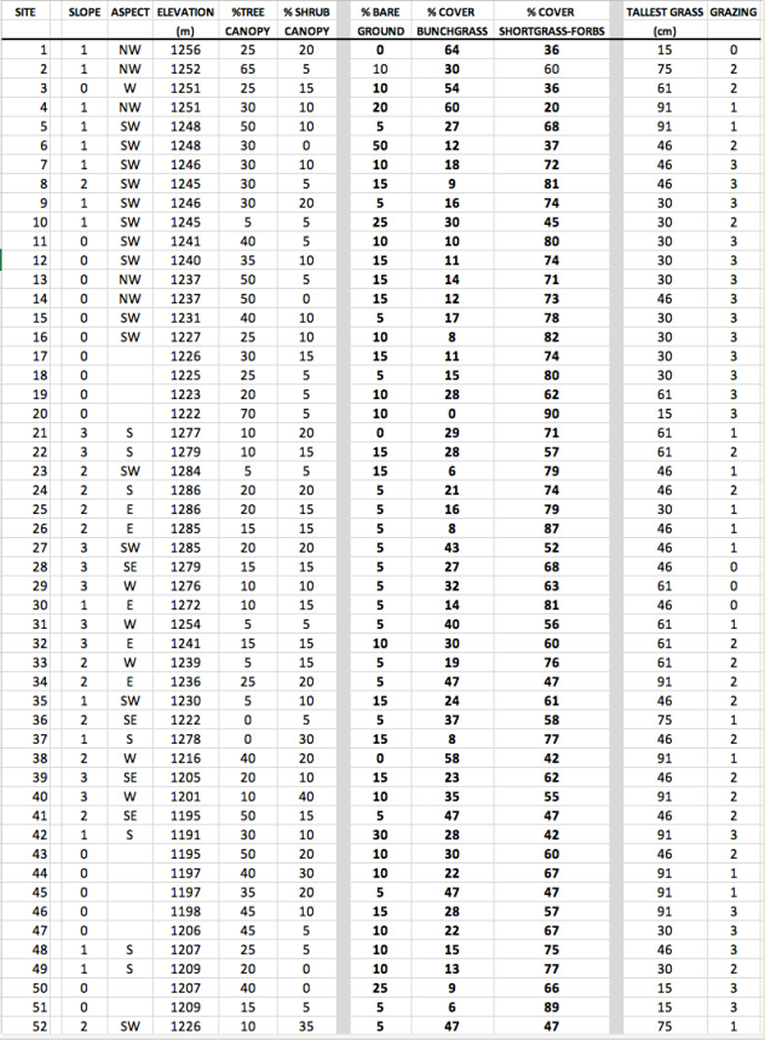
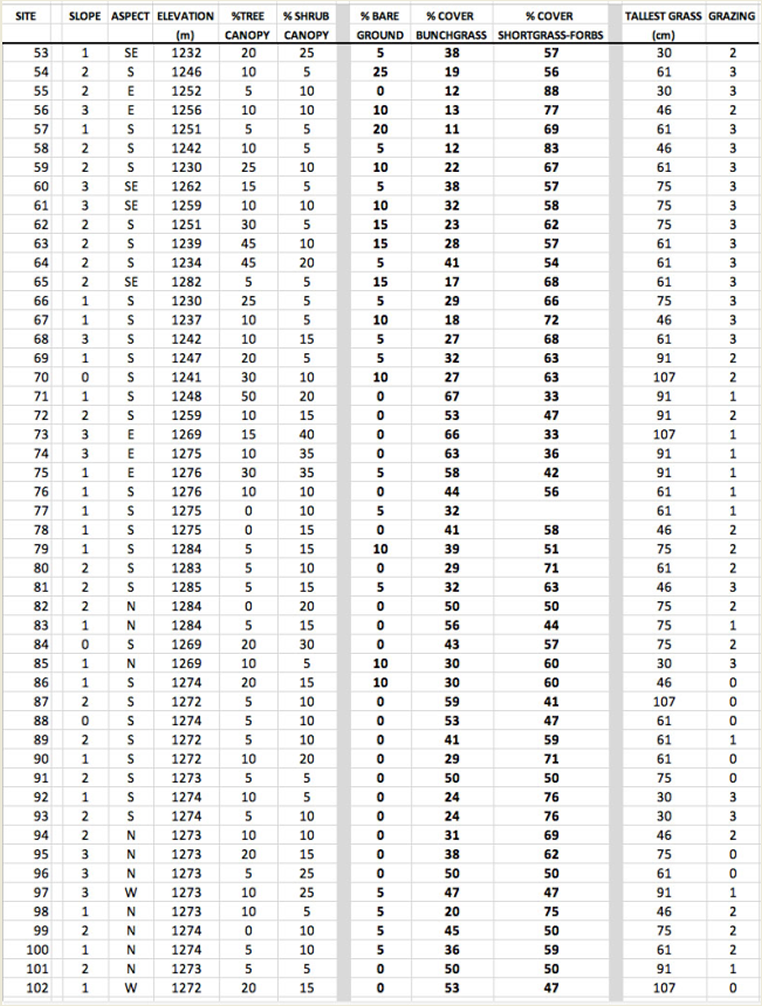
In a second part of the survey, hereafter referred to as “phase 2”, the author surveyed 102 sites over approximately 4.8 km of development road where no Botteri’s Sparrows had been detected in either survey winter. Starting and ending points were at least 180 m from any Botteri’s detections over the past two winters. These surveys were done on foot, stopping every 100 paces (~90 m) and subjectively evaluating a 20 m radius circle, generally half on each side of the road, or, where the slope direction diverged on opposite sides in a circle on one side, and alternating sides at each stop. Mowed yards and houses were bypassed without survey. At each stop the elevation was measured, slope exposure direction determined and the degree of slope was subjectively estimated. In addition, a general assessment was made of the percent trees and shrubs canopy, percent coverage by open ground, “bunchgrasses” and short grasses and forbs, and the tallest grasses measured. Lastly, an estimation was made as to the degree of grazing, using the same subjective standards used in phase 1. The author followed as closely as possible the protocol and standards utilized in the Botteri’s Sparrow site survey (see previous section) except that plants were not identified to species and density was not estimated.
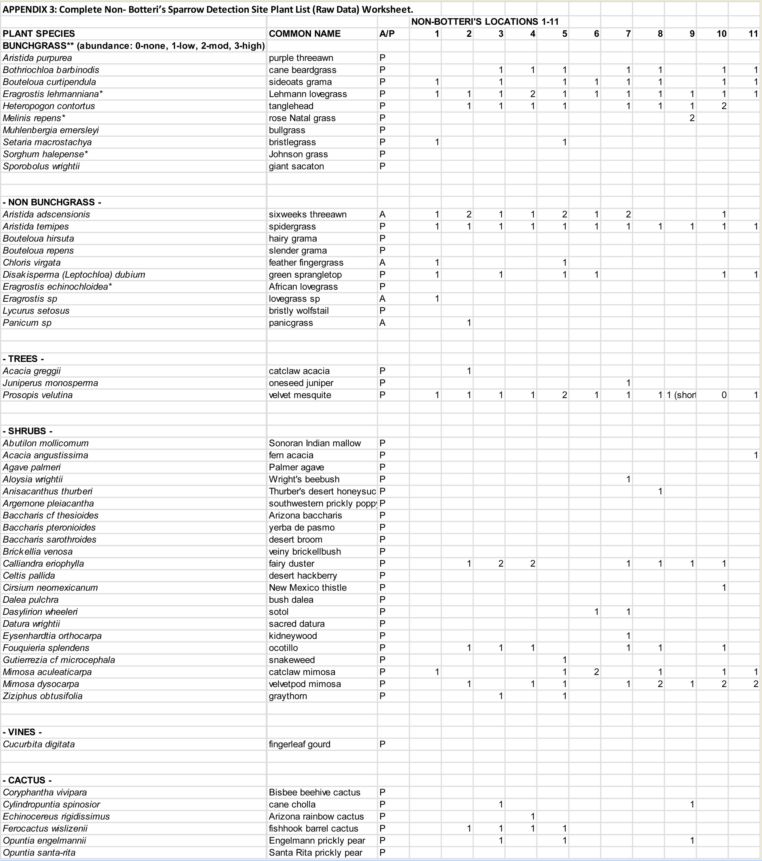
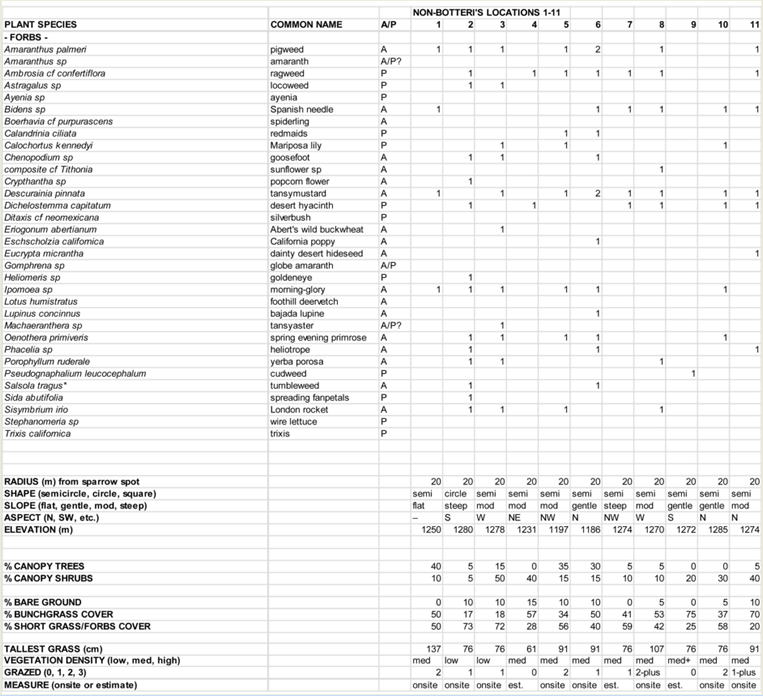
In the course of the phase 2 survey, 11 locations that superficially matched aspects of the Botteri’s Sparrows detection locations (i.e., relatively un-grazed areas with high percent of bunchgrasses and brushy cover), but where none had been detected, were selected to do complete vegetative analysis. On 13 February, this “phase 3” of the study looked more carefully at selected non-Botteri’s Sparrow detection locations for vegetative cover. This was conducted by Carnahan and the author, using the same protocol as used at the site surveys.
RESULTS
 |
| Figure 5: Typical habitat, as seen from the junction of the paved end of Circulo Montana and Patagonia Lake Rd. The gully drains west. Note the overall greater variety and density of vegetation on the left (generally north-facing) compared with the right (south-facing) slope. Survey bird “J” responded here from the shrub with the red dot. Two birds were found in this spot in the 2013 winter survey. Cover “density” was judged to be “medium” here. |
 |
| Figure 6: Site “E”. Note thick and rather tall north-facing ground cover. The “density” of cover was judged to be “high” at this location. In the winter of 2013 there was a bird here also. |
 |
| Figure 7: The tallest bunchgrass was this giant sacaton (Sporobolus wrightii) at site “N”. The bird responded to playback at about a 3 m distance by chipping, hidden within the sacaton base, then finally flew to the shrub to the right of center. This and two others immediately near it are the only sacatons apparent in the survey area. |
The two main roads of Lake Patagonia Ranch Estates follow ridge tops, and are generally oriented northwest to southeast, and draining south-west (see Figure 5). Side roads generally leave the ridge lines and end in the drainages. The very limited flat terrain is found in the form of alluvial benches and flood plains, mostly along the north section of Circulo Montana. Elevations of all the roads surveyed varied from 1191 to 1323 m; range: 132 m). Elevations of Botteri’s Sparrow detections varied from 1259 to 1315 m (median: 1284.5 m). In the phase 2 (non-Botteri’s Sparrow detection sites) survey the elevations ranged from 1191 to 1286 m (median: 1271 m). Of interest is that all of the Botteri’s Sparrows were detected in the upper 50th percentile of the local altitude range, and that 57% of these phase 2 sites surveyed were in the lower half of that altitude range. The steepness of the slope at Botteri’s Sparrow sites was generally gentle to moderate; phase 2 sites similar, with perhaps a few more that were notably steep. Botteri’s Sparrow detections were made 64% of the time on northeast to northwest exposures. In contrast, the phase 2 sites were 62% southeast to southwest exposures. At the 4 detection locations on southeast to southwest-facing slopes there was ambiguity because each site was somewhat flat, and 3 of the 4 birds appeared from nearby multi-directional-sloped gullies with at least some north slope and its associated vegetative cover (Table 1).
All detection sites had at least one velvet mesquite (Prosopis velutina) and a variety of shrubs, predominantly mimosa (Mimosa spp.) and fairy duster (Calliandra eriophylla), that covered about 15% of the typical site. At 79% of the detection sites the overall vegetation cover was subjectively judged as “moderate”, the median tallest grass height was 1.14m, and the degree of grazing was judged as none to light at 93% of the sites. Most of the predominant species of grasses were native, with the exception of Lehmann lovegrass (Eragrostis lehmanniana), and in general the percent “bunchgrass” cover exceeded the percent shortgrasses and forbs cover by about 20 percent (median ratios of 53% to 44%). The site illustrated in Figure 5 was typical, that in Figure 6 was the most densely vegetated and that in Figure 7 was unusual in having the only three giant sacatons (Sporobolus wrightii) observed on any of the survey routes. The most common plant associations at the 2014 detection sites are found in Table 2. A complete plant list by site is found in Appendix 1.
Most phase 2 sites (non-Botteri’s Sparrow detection sites) had at least a few of the same species of trees and shrubs. The mix of grasses at these sites, however, was quite different, as were range conditions. The median percent suface cover of “bunchgrasses” to short-grasses and forbs was 30% to 63%, with median tallest grass far shorter, at only 0.61 m). In addition, these sites generally showed greater signs of recent grazing, with 66% of the survey sites moderately to heavily grazed, compared with only one of 14 Botteri’s Sparrow sites showing even moderate grazing signs. Survey points along the north end of Circulo Montana from Patagonia Lake Road and for about 1.6 km, for example, are in a rather flat alluvial bench/flood plain with 30% mesquite (Prosopis sp) canopy, with only about 15% “bunchgrasses” compared to about 74% short grasses and forbs, and with 72% of all the grasses heavily grazed. The south (paved end) of Circulo Montana from the top of Javalina Court to the end of the pavement, and in addition, south facing side streets Escalone Court, Javalina Court and Guajalote Court, where it is rather uniformly steep, had only 22% “bunchgrasses” compared with 59% short grasses and forbs, and with 73% heavily grazed (Fig. 8). The south side of the far end of Lado de Loma Road similarly tended to be mostly steep and had only a few tufts of bunchgrasses (summary in Table 1; complete data in Appendix 2).
The selection of sites from phase 2 for the more in-depth phase 3 site surveys was difficult and somewhat arbitrary, but an attempt was made to subjectively select locations that superficially resembled the density and superficial vegetation of that of Botteri’s Sparrow detection sites, and to spread these sites geographically. These sites are shown on the linked map with a large letter “P”; see here.
The general data summary from these phase 3 site surveys appears in Table1. The differences between Botteri’s Sparrow detection sites and these samples are not dramatic, but one notices that some of these sites (36%) are at lower elevation than any Botteri’s Sparrow sites. In addition, vegetative species are similar, but the average tallest grasses are shorter and the degree of grazing appeared to be markedly greater. A complete plant list is found in Appendix 3
 |
| Figure 8: Typical south-facing slopes along paved end of Circulo Montana, showing lack of any good habitat for Botteri’s Sparrows due to southern exposure and grazing. Overall “density” of cover would be judged to be “none” to “low” for most of the central area here. |
Table 1: Site descriptions and overall mix of vegetation covers at survey sites; subjectively determined by Carnahan and Schmierer. Bunchgrasses were defined as perennial grasses forming thick “tufts” or “tussocks” rather than small tufts or mat-like ground cover. |
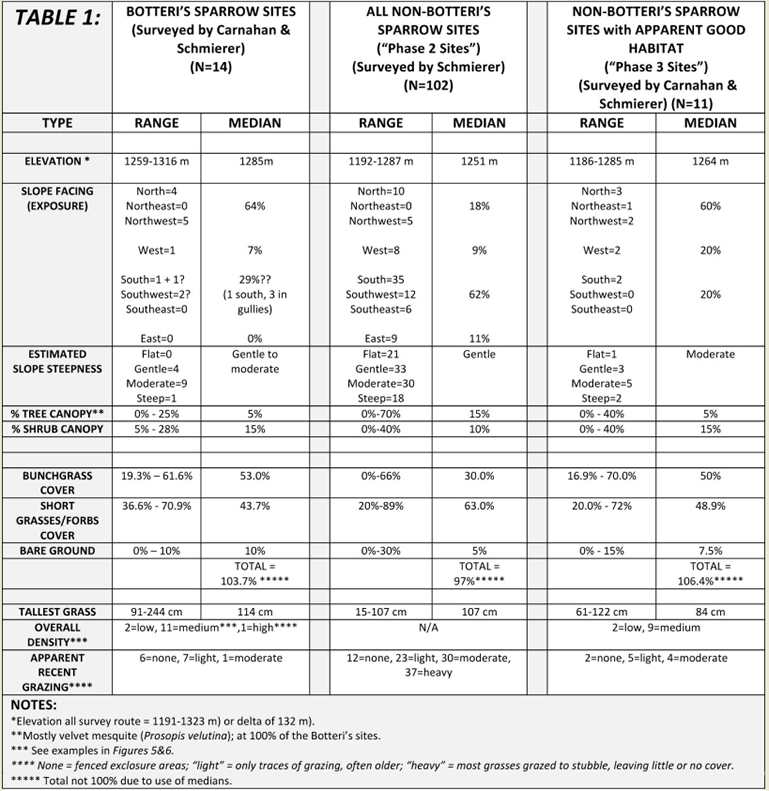 |
Table 2: List of Vegetation found in at least 33% of the Botteri’s Sparrow detection sites; including phase 3 surveyed plants for comparison. Those highlighted with gray are plants with seeds known to be consumed by at least some species of sparrows. (Complete plant lists available at Appendix 1 and 2) |
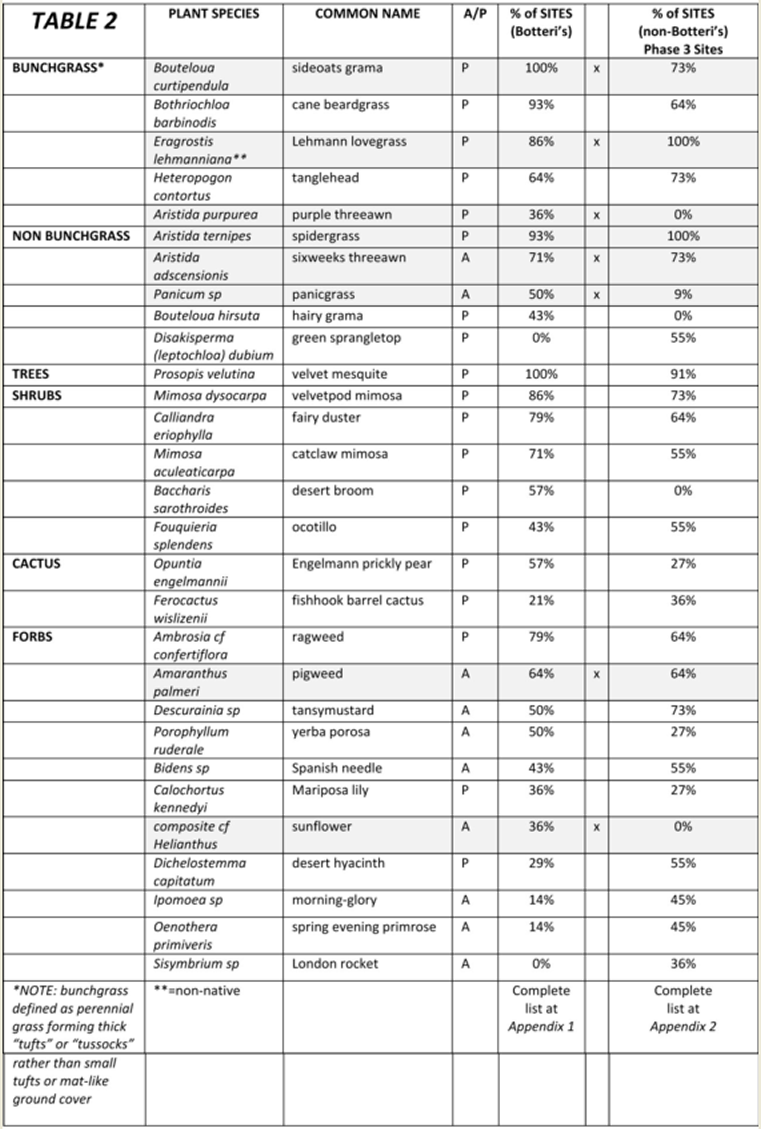 |
DISCUSSION: RELATIONSHIPS BETWEEN BOTTERI’S SPARROWS AND
STUDIED HABITAT
The Lake Patagonia Ranch Estates development includes lots from about 2 to 16 hectares in size. Most lots are undeveloped and fewer than ten percent are fenced to exclude cattle. The vegetative surveys suggest that most of the area could best be described as being between “native” and “exotic” grasslands; Lehmann lovegrass is present throughout the area, but nowhere is it dominant or exclusive. The selective grazing by livestock encourages the spread of lovegrass, which is not a preferred food source for them (Webb and Jones 2012). Without prior data it is difficult to assess where the surveyed area stands in any evolution toward true exotic grassland status. Through open-range laws variable cattle grazing occurs, and several individuals graze horses at large. The heaviest grazing pressures are at the southeast end of the development and in the alluvial floodplains along much of Circulo Montana.
The only reference regarding wintering Botteri’ Sparrows in the online Birds of North America account is that they winter in “... grassy washes, weedy fields and oak woodlands with tall grass understory.” (Webb 1985; Russell and Monson 1998, in Webb and Bock 2012)
The breeding success of Botteri’s Sparrows appears to be more directly linked to the structure of the habitat, the height and density of the cover, than the actual floristics, perhaps by providing the cover necessary for the survival of young (Jones and Bock 2005). These authors further suggest that the incursion of Lehmann lovegrass into native grasslands, even when un-grazed, augments the potential Botteri’s Sparrow breeding habitat due to its height and density (Jones and Bock 2005). As it is not readily grazed by livestock its persistence into winter is encouraged, providing Botteri’s Sparrows with an alternate grassland habitat to its preferred sacaton, and one that perhaps will endure all but the heaviest grazing pressures.
In the current study, comparing selected sites where winter Botteri’s Sparrows have been detected with sites where they have not been detected, likewise suggests that the general density of ground-cover, characterized by medium to tall “bunchgrasses” and low shrubs, may be an important factor in providing a favorable winter habitat. These are found locally more on north-facing slopes and gullies, and generally show moderate or less grazing. Although not actually measured in the study, it was noted that the cover at several of the sites was enhanced by windblown spidergrass (Aristida ternipes) in matted drifts against shrubs and taller “bunchgrass” clumps, creating an even more dense cover. The reason for the dense cover preference in winter is less clear than has been theorized for breeding birds, however, as it cannot be related directly to the survival of young. It may be that dense cover is the species’ regular winter habitat and wintering birds are sedentary post/pre-breeders. Alternately or in addition, this habitat may offer protection from predators, winter cold, and wind, thus helping birds conserve energy in addition to providing food.
Food preferences during breeding season have been well documented. They consist of predominantly animal foods (70% to 86%), mostly Orthoptera, Lepidoptera and various larvae, with the remainder mostly seeds (Webb and Bock 2012). During winter, however, when nighttime temperatures may reach well below freezing, energy requirements must be high but availability of animal matter is presumably minimal. The only direct references to Botteri’s Sparrow winter diet are to a single bird’s stomach content that was reported to contain seed and grit (Wolf 1977 in Webb and Bock 2012), and “...seeds of chickweed (Stellaria media), plantain (Plantago sp.), wood sorrel (Oxalis sp.), sedge (Carex spp.), panic grass (Panicum spp.) and other grasses, and sorghum (Sorghum sp.)...” (Webb and Bock 2012). Of these cited potential food sources, panic grass was present at half of the Botteri’s Sparrow sites and only one non-Botteri’s Sparrow site, and Johnson grass (Sorghum halepense) was found at three detection sites and but no no-detection site.
Other wintering southwestern sparrow species have been studied in detail. Over 65 species of seeds were found in the crops of 16 species of sparrows in winter in southwestern New Mexico (Desmon et al 2008). Seed utilization appears to be complex, with the species of bird, the size of the species, the bill size, the quantity of seed in the seed bank, the nutritional value of the seed, the ease of eating the seed (whether it could be eaten whole or needed to be hulled), the protective devices inherent to each seed, etc., all being factors (Desmon et al 2008; Mendez-Gonzalez 2010). Despite a seed bank of over 90 species, five seed species accounted for 80% of seeds consumed in the species studied (Desmon et al 2008; Mendez-Gonzalez 2010): sand and spike dropseed (Sporobolus spp.), perennial grasses, found at only 7% of the current study Botteri’s Sparrow sites, and none at the studied non-Botteri’s Sparrow sites; feather fingergrass (Chloris virgata), an annual grass, found at no Botteri’s and only one studied non-Botteri’s site; stinkgrass (Eragrostis cilianensis), an annual grass, found at 86% and 100% of sites respectively; amaranth (Amaranthus spp.), an annual forb: found at 64% of both types of sites; and carpetweed (Mollugo verticillata), an annual forb, not found at any survey site. Amaranthus seeds are high in fat and overall energy (Mendez-Gonzales 2010).
Although there is conflicting information, several studies have reported positive relationships between winter sparrow abundance and seed abundance, suggesting sparrows cue in on overall seed production (Pulliam and Parker1979, Dunning and Brown 1982, Grzybowski1983, Ginter and Desmond 2005 in Desmon et al. 2008). This in turn may influence seed availability in a small area such as the survey routes covered in the current research. The annual rainfall in the study area averages about 41 cm, but can be quite variable in both extent and local event coverage. For example, the 2012 monsoonal rainfall was 38.6 cm but in 2013 was only 26.0 cm (measured at one location within 2 km of any Botteri’s Sparrow detection in this study [personal data]).
Assuming that good monsoonal rain produces an abundance of seeds, however, is not necessarily a predictor of greater number of species or individuals utilizing those seeds (Whitmore 1979; Desmon et al. 2008). Further complications in interpreting any data emerge from unknown factors such as whether the winter range or territory of the Botteri’s Sparrows are the same as they maintain in breeding season, how strong the territory might be, whether they are mobile in tracking seed resources and precisely which local seeds are preferred even when seeds are available in abundance.
SUMMARY AND CONCLUSIONS
The winter 2013-14 marked at least the fourth consecutive winter season that significant numbers of Botteri’s Sparrows were present in the Lake Patagonia Ranch Estates area. Although the numbers detected were lower than during the previous winter this adds to the evidence that this might be a permanent pattern of winter range extension and not an irruptive event. The reason that the number of detections this year was only 60% of last year, despite a 10% increase in hours of effort by the author, is not clear. It may represent real change or represent a less attractive seed crop, perhaps secondary to lower monsoonal rains. Alternately, survey routes do not necessarily bisect all the best potential habitats, and so year to year differences may represent variations in detections of this rather secretive species.
Botteri’s Sparrows are a regular breeding species in the study area. Their winter territorial behavior remains largely unknown. The personal observation that birds were re-findable at the same locations over a period of at least eight weeks, and the fact that almost half of the birds were detected less than about 90 m from locations where they were detected in the winter of 2013 might suggest some form of sedentary behavior or territorial fidelity. The fact that the related Cassin’s Sparrows shows sedentary behavior supports this hypothesis. Without more definitive evidence, however, we cannot rule out that these birds were merely seeking the most favorable habitat.
The aggressive response to playback perhaps offered the most convincing evidence that some territory was in fact being defended. It is unclear from our survey and subsequent observations whether the relative strength of the aggressive behavior and chipping calls were elicited from one or both sexes, whether they were from wintering or resident bird “reserving” a breeding territory, or whether the reaction strength was merely due to the position of the intruding playback within that potential territory. The trilled calls by several agitated birds may represent a unique winter territory call or may be similar to breeding defensive calls, unfamiliar to the author.
Comparing the habitats where Botteri’s Sparrows were detected in these surveys with habitats where the author found no wintering birds, the differences appeared mostly in the fact that sparrow detection sites had taller and denser ground cover. This density was evident mostly in the form of greater percent “bunchgrasses” cover plus a relatively lower degree of grazing. Most birds were detected on north-facing slopes and on the edge of drainage gullies, in fenced livestock exclusion areas, or where grazing was minimal, all of which would support the before-mentioned density-of-cover factor. This is consistent with the birds’ breeding habitat preferences as expressed in the previously cited literature. There is evidence that the related Cassin’s Sparrows in this area are in fact affected negatively by grazing (Ruth 2000).
Although the number of Botteri’s and selected non-Botteri’s Sparrow study sites was small, no notable differences in actual floristics were found. Most of the common seed species that could be documented in the literature as being preferred as winter food by sparrows, if found at all, were found at both Botteri’s and the selected non-Botteri’s Sparrow survey sites. The fact that all the birds were detected in the top 50th percentile of the available elevations of the area may have significance. By personal observation the winter nighttime temperatures in the lower elevations of the development are several degrees (2-3 degrees C) colder than the ridge tops. In turn the persistence of flowers in the fall and the emergence and budding of early spring plants differs notably between ridge-tops and lower drainages, perhaps encouraging the late persistence and early emergence of potential animal food sources on both a daily and seasonal basis.
FURTHER STUDY
As with any research, perhaps, this survey has probably raised more issues than it has solved. The following future steps would be useful:
- The author plans to continue to monitor the local rainfall to assess whether this may be a factor in the number of wintering birds present during the following winter.
- Recordings of the winter trilled call should be attempted.
- Capture and banding would establish a base to identify the sex of wintering birds, and, in addition, possibly information about breeding-winter site fidelity and winter territory size. (Impediments to this might present themselves because of private land with numerous landowners.)
- It is the author’s opinion that reporting birds, especially those of special interest, such as Botteri’s Sparrows in fall, winter and early spring, into the eBird data base is extraordinarily important for future tracking and research regarding this species. If all birders use this mechanism, it then becomes an instant and powerful research source to track this and other species.
ADDENDUM
SUMMARY of 17, 18 and 19 JANUARY 2015 SURVEY:
A follow-up survey was done by the author on 17-19 January 2015. The protocol was generally the same as that used in the 2013 and 2014 surveys, with an effort of 19.8 km (11.8 km on foot and 8.0 km by car) over 13 hours, plus a re-survey of 0.5 km of Circulo Montana on 21 January. A total of 39 birds were detected at 35 locations, compared with 25 birds at 20 locations in 2013 and 15 birds at 12 locations in 2014. Thirty-seven (95%) of the birds were found in the upper 50th percentile of the elevation differential between the highest and lowest elevations in the surveyed area. 2014 monsoon rainfall in the area was 41.7cm.
ACKNOWLEDGMENTS
Thanks to Pierre Deviche for his encouragement, suggestions and editorial assistance; to the peer-reviewers for their important comments; and a special thanks to Sue Carnahan who cheerfully took many hours from her busy life to work with the author on this project.
LITERATURE CITED
Arcese, P., M.K. Sogge, A.B. Marr and M.A. Patten. 2002. Song Sparrow (Melospiza melodia), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/704doi:10.2173/bna.704
Agudelo, M.S., M.J. Desmond and L. Murray. 2008. Influence of desertification on site occupancy by grassland and shrubland birds during the non-breeding period in the northern Chihuahuan Desert. In Birds of the US-Mexico borderlands: distribution, ecology and conservation. J. Ruth, T. Brush and D. Krueper (assoc. editors). Studies in Avian Biology 37: 84–100. A Publication of the Cooper Ornithological Society. http://sora.unm.edu/sites/default/files/journals/sab/sab_037.pdf
Avibase – the world bird database. http://avibase.bsc-eoc.org/avibase.jsp (Searched on 28 January 2014).
Desmon, M. J., C. Mendez-Gonzales and L. B. Abbott. 2008. Winter diets and seed selection of granivorous birds in southwestern New Mexico. In Birds of the US-Mexico borderlands: distribution, ecology and conservation. J. Ruth, T. Brush and D. Krueper (associated editors). Studies of Avian Biology No. 37: 101–112). A Publication of the Cooper Ornithological Society.
Dunning, Jr., J.B., R. K. Bowers, Jr., S.J. Suter and C.E. Bock.1999. Cassin’s Sparrow (Peucaea cassinii), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online: http://bna.birds.cornell.edu/bna/species/471doi:10.2173/bna.471
eBird 2014. An online database of bird distribution and abundance [web application]. eBird, Ithaca, New York. Available: http://www.ebird.org. (Accessed: 28 January 2014)
Howell, S. N. G. and S. Webb. 1995. The Birds of Mexico and northern Central America. Oxford Univ. Press, Oxford.
Jones, Z.F. and C.E. Bock. 2005. The Botteri’s Sparrow and exotic Arizona grasslands: an ecological trap or habitat regained? Condor 107: 731-741. Cooper Ornithological Society. http://www.bioone.org/doi/full/10.1650/7741.1
Leahy, C. 1982. The birdwatcher’s companion: an encyclopedic handbook of North American birdlife. Hill and Wang, New York.
Mendez-Gonzalez, C.E., Ph.D. 2010. Influence of seed resources on the diet, seed selection, and community dynamics of wintering birds in semi-arid grasslands. New Mexico State University, 112 pages; 3534178
http://gradworks.umi.com/35/34/3534178.html
Rising, J.D. 1996. A guide to the identification and natural history of the sparrows of the United States and Canada. Academic Press, Inc., San Diego.
Russell, S. M. and G. Monson. 1998. The Birds of Sonora. Univ. of Arizona Press, Tucson.
Ruth, J.M. 2000. Cassin’s Sparrow (Aimophila cassinii) status assessment and conservation plan. Biological Technical Publication BTP-R6002-1999. U.S. Department of the Interior, Fish and Wildlife Service, Denver, CO http://www.fort.usgs.gov/Products/Publications/556/556.pdf
Schmierer, A. 2013. Current post-breeding, winter, and spring status of Botteri’s Sparrow (Peucaea botterii arizonae) in Arizona. Arizona Birds Online: the journal of the Arizona Field Ornithologists.
http://www.azfo.org/journal/documents/BOSP_Article_14Sep2013_Final.pdf
Garrigues, R. Recorded on 2 May 2010 in Costa Rica. From The Internet IBC Bird Collection: http://ibc.lynxeds.com/ (Accessed 28 January 2014)
Webb, E.A. and C.E. Bock. 2012. Botteri’s Sparrow (Peucaea botterii), The Birds of North America Online (A. Poole, Ed.). Ithaca: Cornell Lab of Ornithology; Retrieved from the Birds of North America Online
http://bna.birds.cornell.edu/bna/species/216doi:10.2173/bna.216
Whitmore, R.C. 1979. Temporal Variation in the Selected Habitats of a Guild of Grassland Sparrows. The Wilson Bulletin Vol. 91 :592-598 Published by: Wilson Ornithological Society Article Stable URL:
http://www.jstor.org/stable/4161273
 |
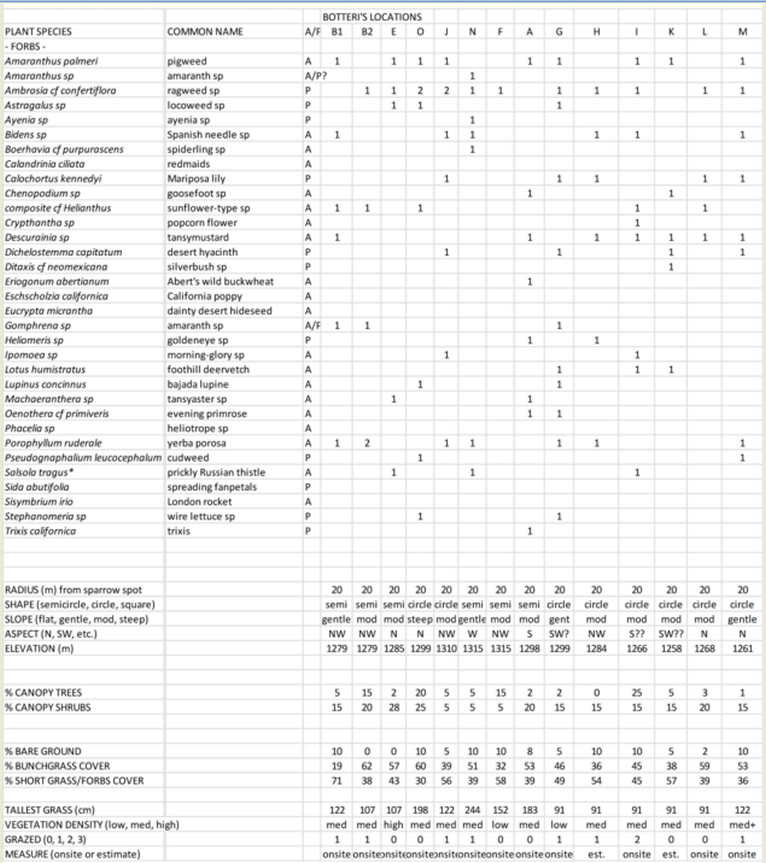 |
| APPENDIX 2: Phase 2 survey of sections of road where no detections had been made in either year. |
 |
 |
 |
 |
|
©2005 |
HOME | | | REPORT SIGHTINGS | | | PHOTOS | | | BIRDING | | | JOURNAL | | | ABOUT US | | | CHECKLISTS | | | AZ BIRD COMMITTEE | | | EVENTS | | | LINKS |